Feldmann, F.*, Silva Jr., J.P.+ & A.V.R. Jayaratne#
*Institut für Pflanzenkultur, Solkau, Deutschland; +Centro Nacional de Pesquisa Agroflorestal da Amazonia, Manaus, Brasilien; #Rubber Research Institute of Sri Lanka, Agalawatta, Sri Lanka
Nutzung der
arbuskulären Mykorrhiza in Baumschulen der Tropen am Beispiel des
Kautschukbaumes Hevea spp.
Utilization of arbuscular mycorrhiza in tropical
nurseries exemplarily demonstrated for the rubber tree Hevea spp.
The experimental introduction of arbuscular mycorrhizal fungi into the plant production process of the rubber tree (Hevea spp.) pointed out the economic importance of the mycorrhizal technology as an efficient mangagement factor in practice. Better growth of young plants, shorter growth period in nurseries, decreased plant losses in spite of the presence of root and leaf pathogens, reduction of pesticide use, higher success in budgrafting, reduced losses after planting in the field, and finally better growth in the field with the consequence of earlier crown-budding are the economically positive valuating factors.
Zahlreiche der ein- und mehrjährigen tropischen und subtropischen
Nutzpflanzen werden von arbuskulären Mykorrhizapilzen besiedelt und viele sind
von der Ausbildung der Symbiosen mehr oder weniger stark abhängig (Janos, 1987). Das natürliche Vorkommen
von arbuskulären Mykorrhizapilzen (AMP) wird bei der Anlage von Nutzflächen
durch die begleitend angewendeten Maßnahmen stark beeinträchtigt (Feldmann und Lieberei, 1994) und führt dazu, daß eine ausreichende
Versorgung ausgepflanzter Nutzpflanzen mit geeigneten AMP im Feld oft nicht
gewährleistet ist (Feldmann et
al. 1995a). Pflanzenbaumaßnahmen, die auf eine Verbesserung der ökologischen
Situation von Anbauflächen in den Tropen gerichtet sind (z.B. Mischkulturanbau
und weitgehende Belassung von Sekundärvegetation), erwiesen sich für den Aufbau
wirksamer, autochtoner AMP-Populationen nützlich (Feldmann et al., 1995b). Bevor die Nutzpflanzen ins Gelände
gepflanzt werden, durchlaufen sie ihre frühe Entwicklungsphase in Baumschulen,
wo - vergleichbar zu der Produktion von Jungpflanzen in den gemäßigten Breiten
- vorab sterilisierte Pflanzsubstrate zum Einsatz kommen. Bei der
Sterilisierung gehen im Substrat befindliche AMP zugrunde, so daß die
Jungpflanzen ohne Inokulation geeigneter Symbionten nicht zur Ausbildung der
für sie essentiellen Lebensgemeinschaft gelangen können.
Der Kautschukbaum Hevea brasiliensis ist
eine Weltwirtschaftspflanze mit großer Bedeutung für die Volkswirtschaft
verschiedener Länder in den Tropen. In manchen Anbauländern wird der Feldanbau
durch Krankheiten der Pflanze sehr stark beeinträchtigt. In Brasilien führt das
Auftreten von Blattkrankheiten, insbesondere der Südamerikanischen
Blattfallkrankheit, hervorgerufen durch Microcyclus
ulei, dazu, daß ein nachhaltiger Plantagenanbau des Kautschukbaumes in
nennenswertem Umfang in der Herkunftsregion Amazonien bislang nicht möglich ist
(Junqueira et al., 1988). Brasilien
insgesamt ist sogar von der Einfuhr von Kautschuk abhängig. In Sri Lanka
dagegen ist der Anbau von Hevea vor
allem durch Wurzelkrankheiten, z.B. die Weissfäule, hervorgerufen durch Rigidoporus lignosus, bedroht (Jayaratne, 1982).
Zwischen 1987 und 1998 studierten wir deshalb in mehreren
Forschungsprojekten die Wirkung der Mykorrhizierung von
Kautschukbaum-Jungpflanzen auf ihr Resistenz- bzw. Toleranzverhalten gegenüber
Blatt- und Wurzelkrankheiten, ihre Biomasseentwicklung, ihren Ernährungsstatus
und den Propfungserfolg unter dem Einfluss der Symbiosen und ihre
Stresstoleranz während der Produktion in der Baumschule und unmittelbar nach
dem Auspflanzen ins Feld. Neben diesem intensiven Studium des Kautschukbaumes
wurden im gleichen Zeitraum zahlreiche andere tropische und subtropische
Nutzpflanzen auf ihre Mykorrhizierfähigkeit untersucht, um weitere potentielle
Zielpflanzen und Einsatzbereiche abschätzen zu können.
Orte der Durchführung der Experimente waren das Centro Nacional de Pesquisa
Agroflorestal da Amazonia, Manaus, Brasilien, und das Rubber Research Institute
of Sri Lanka, Agalawatta, Sri Lanka. Die Parameter der Pflanzenkultur und
Analysemethodik zum Studium der Sämlingsentwicklung und Ernährung der Pflanzen,
die Studien zur Blattresistenz und der Wurzeltoleranz gegenüber Fusarium solani, einem Schaderreger, der
bei Hevea zu Wachstumsreduktionen und
Blattwelke (Liyanage und Dantanarayana, 1983), aber selten zu
Pflanzenausfällen führt, sind bei Feldmann (1991) dargestellt und fanden in Brasilien statt. Als
Mykorrhizapilze wurden hier verschiedene Linien von Glomus etunicatum eingesetzt. Das Masseninokulum wurde nach Feldmann und Idczak (1994) produziert.
Alle übrigen Versuche fanden in Sri Lanka statt. Wesentliche Unterschiede
zwischen dem Versuchsaufbau in beiden Ländern lagen im pH-Wert der Substrate
(beides lehmige Laterite mit starkem Phosphatmangel): in Brasilien pH 4,8, in
Sri Lanka pH 6,2. In Sri Lanka wurden Linien der AMP Gigaspora margarita, Acaulospora spec. und eine aus einer Plantage
isolierte Mischung autochtoner Pilze, vermehrt in identischen Bodensubstrat,
eingesetzt. In beiden Ländern wurden unterschiedliche Kautschukbaum-Klone
verwendet: In Brasilien Unterlagen von maternal definierten Fx 25 - Samen,
Edelreiser der Klone Fx 4098, Fx 3925, RRIM 600; in Sri Lanka wurden maternal
definierte Sämlinge des Klones PB 86 bzw. Jungpflanzen der Klone RRIC 100
getestet.
Die Methodik der Mykorrhizaanalyse ist bei Feldmann et al. (1999, in diesem Band) dargestellt. Alle
übrigen relevanten Daten sind in den Legenden der Tabellen und Abbildungen
wiedergegeben.
Wurzelproben der Nutzpflanzen aus Tab. 3 wurden im Freiland, in Botanischen
Gärten oder Pflanzenproduktionsbetrieben gesammelt, zahlreiche wurden als
Saatgut oder Stecklinge mit AMP inokuliert und ihre Mykorrhizierfähigkeit nach
Wachstum im Gewächshaus festgestellt.
Nach Auskeimen der Samen erfolgte eine Besiedelung der
Kautschukbaum-Sämlinge erst mit Eintreten einer Netto-Photosynthese-Leistung
der ersten Laubblätter. In den ersten 210 Tagen nach Inokulation wurde das
Wurzelsystem zu 54% ± 12% besiedelt. In dieser Zeit veränderte sich das Wachstum der mykorrhizierten
Pflanzen gegenüber der nicht-inokulierten Kontrollvariante. Es kam zu einer
verstärkten Biomassebildung mykorrhizierter Pflanzen, nachweisbar auf der Ebene
zahlreicher Wachstumsparameter (Abb. 1). Für die Pflanzenproduktion besonders
bedeutsam erwies sich eine Vergrößerung der Pflanzenhöhe in Verbindung mit
einem stärkeren Durchmesser der Pflanzen. Dadurch gelangten die Jungpflanzen
sehr viel früher in das Entwicklungsstadium, das für die Pfropfung der
Edelreise erlangt werden muß. Gleichzeitig vermochten die mykorrhizierten
Jungpflanzen dem Substrat mehr Nährstoffe zu entziehen (Abb.1). Der Gehalt an
Nährstoffen in den Pflanzengeweben war nicht signifikant erhöht; vielmehr
setzten die Pflanzen die vermehrt aufgenommenen Nährstoffe in erhöhte Wachstumsleistung
um.
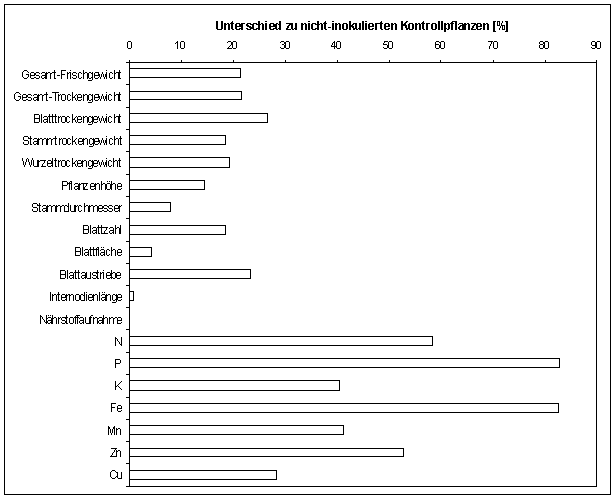
Abb.
1: Verstärktes Wachstum und
verbesserte Ernährung von Kautschukbaum-Sämlingen unter dem Einfluß der
arbuskulären Mykorrhiza (Glomus etunicatum)
Für die Untersuchungen wurden 180 Tage alte Sämlinge des Kautschukbaumes
verwendet. Die Anzahl der untersuchten Pflanzen lag bei mindestens 30 pro
Experiment in mindestens drei Wiederholungen. Alle dargestellten Werte über 3%
sind signifikant von Kontrollpflanzenwerten unterschiedlich (Kruskal-Wallis
One-way ANOVA; 95% Intervall)
In einem weiteren Experiment wurde sowohl der Durchmesser der Sämlinge als
auch die Pflanzenhöhe von verschiedenen AMP-Linien unterschiedlich beeinflußt
(Tab. 1). Alle drei getesteten AMP-Linien verbesserten das Wachstum signifikant
gegenüber nicht-mykorrhizierten Kontrollpflanzen. Mykorrhizierte Pflanzen
erreichten nicht nur die Propfreife früher, sondern zeigten zudem im Falle der
Inokulation mit Gigaspora margarita
eine geringere Verlustrate von Edelreisern als nicht-mykorrhizierte
Propfunterlagen (Tab. 1).
Tab. 1: Spezifische Wirksamkeit verschiedener AMP-Linien auf die
Entwicklung von Propfunterlagen und den Propferfolg
bei Hevea-Jungpflanzen (Klon Pb 86) in tropischen
Baumschulen acht Monate nach Auskeimen und gleichzeitiger Inokulation.
Werte in Spalten mit dem gleichen Buchstaben sind statistisch nicht signifikant unterschiedlich; LSD 0,05%, 5 Wiederholungen mit 10 Pflanzen)
|
AMP-Linie |
Pflanzenhöhe [cm] |
Stammdurchmesser [mm] (12cm über Boden) |
Propferfolg [%] |
|
ohne (Kontrolle) |
81,1 c |
69 c |
81,8 b |
|
Gigaspora margarita |
101,9 a |
90 a |
93,0 a |
|
Acaulospora spp. |
88,9 b |
79 b |
83,4 b |
|
Autochtoner AMP |
86,3 b |
76 b |
84,2 b |
Die Mykorrhizasymbiose führte bei drei Kautschukbaumklonen zu einer erhöhten
Resistenz gegenüber dem Blattpathogen Microcyclus
ulei. Die Verzögerung der Pathogenentwicklung im Blatt mykorrhizierter
Pflanzen führte zu einer geringeren Größe der Pathogen-Läsionen und zu einer
verminderten Sporulation des Pathogens auf den Blättern (Tab. 2) mit der Folge
nachhaltig veränderten epidemiologischen Verhaltens der Krankheit im
Testpflanzenbestand. Die Wirksamkeit von Glomus
etunicatum war klonspezifisch unterschiedlich, aber in allen drei
pathogen-anfälligen Klonen nachweisbar. Ein vierter, pathogen-resistenter Klon
(IAN 6158) wurde nicht nachteilig durch die Mykorrhizierung beeinflusst.
Kautschukbaumsämlinge, die mit Fusarium
solani in Kontakt gebracht wurden, waren zwar in geringem Umfang durch das
Pathogen befallen, jedoch wurde keine Reduktion des Wachstums beobachtet (Tab.
2). Zu einer vollständigen Verhinderung Pathogen-bedingter
Wachstumsdepressionen kam es an Jungpflanzen, die mit dem Erreger der
Weissfäule, Rigidoporus lignosus,
beimpft wurden. An mykorrhizierten Pflanzen erfolgte hier keine Besiedelung
mehr durch das Pathogen (Tab. 2).
In der Jungpflanzenproduktion des Kautschukbaumes folgt der Phase der
Herstellung von Propfunterlagen aus Sämlingen und der Propfung mit Edelreisern,
die nach Ertragsleistung ausgewählt werden, nach ca. sechs Monaten eine
Transplantation der Baumschulpflanzen an den Bestimmungsort ins Feld. Bei
diesem Umpflanzen gehen z.T. große Mengen an Jungpflanzen durch den
Umpflanzstress verloren. Hinzu kommt, daß die nach der zukünftig zu erwartenden
Latexmenge ausgewählten Klone solange anfällig gegen Microcyclus ulei sind, bis ihre Kronen durch eine erneute Propfung
im Feld gegen resistente Kronen ausgetauscht werden.
Tab. 2: Wechselbeziehungen zwischen Hevea-Klonen und Schaderregern unter dem Einfluß von Mykorrhizasymbiosen
Werte in Zeilen mit gleichen Buchstaben sind nicht signifikant unterschiedlich; Separater Varianz t- Test; 95% Intervall
|
Infizierte Organe |
AMP |
Pathogen |
Parameter |
Hevea-Klon |
ohne AMP |
mit AMP |
|
Blätter |
Glomus etunicatum |
Microcyclus ulei |
Pathogen- sporenzahl |
Fx 4098 |
740a |
80b |
|
|
|
|
[n x 1000/ Blatt] |
Fx3925 |
590a |
120b |
|
|
|
|
|
RRIM 600 |
850a |
580b |
|
|
|
|
|
IAN 6158 |
0a |
0a |
|
Wurzeln |
Glomus etunicatum |
Fusarium solani |
Trockenge-wicht [g] |
Fx 25 |
8,5b |
12,9a |
|
|
Gigaspora margarita |
Rigidoporus lignosus |
Trockenge-wicht [g] |
PB 86 |
6,8b |
7,7a |
Das Auspflanzen mykorrhizierter Jungpflanzen war in unseren Versuchen mit
einer drastischen Reduktion der Verluste verbunden: ohne Mykorrhiza-Inokulation
gingen 25% der Jungpflanzen in den ersten zwei Monaten im Gelände verloren, mit
Inokulation nur noch 3%. Zusätzlich entwickelten sich die überlebenden Pflanzen
im Feld so vorteilhaft, daß zum Zeitpunkt der ersten Kronenpropfung (dieser ist
dann gegeben, wenn ca. 70% aller zu propfenden Pflanzen, einschließlich der
nicht-inokulierten Kontrollpflanzen, eine Höhe von 180cm erreicht haben) mehr
als 2/3 mykorrhizierte Pflanzen gepropft werden konnten. So waren von jeweils
392 Pflanzen bereits 70 % aller AMP-inokulierten Pflanzen für die
Kronenpropfung geeignet, während dies bei der nicht-inokulierten Variante nur
für 30% zutraf.
Jede mykorrhizierfähige Pflanze kann in Abhängigkeit von der jeweiligen
Umwelt und den verfügbaren AMP als Zielpflanze für einen nutzbringenden
Mykorrhizaeinsatz aufgefasst werden. Von 298 untersuchten Nutzpflanzen der
Tropen und Subtropen erwiesen sich 92% (274 Arten) als Wirte arbuskulärer
Mykorrhizasymbiosen (Tab. 3). Die in Tab. 3 nicht aufgeführten Nicht-Wirte
gehörten zu den Familien der Brassicaceae,
Chenopodiaceae und Cyperaceae. Zu
den Wirtspflanzen gehören landwirtschaftlich angebaute Pflanzen, aber auch
solche, die als Zierpflanzen in den gemäßigten Breiten gehandelt und dort im
Gartenbau produziert werden.
Der experimentelle Einsatz von arbuskulären Mykorrhizapilzen in der
Pflanzenproduktion des Kautschukbaumes erwies sich unter Praxisbedingungen als
ökonomisch sehr sinnvolle Pflanzenbaumaßnahme. Besseres Wachstum der
Jungpflanzen und damit kürzere Standzeiten in
Tab. 3: Wirte arbuskulärer Mykorrhizapilze unter wichtigen
Nutzpflanzen der Tropen
und Subtropen
Wurzelproben der aufgeführten Pflanzenarten wurden als mykorrhiziert betrachtet, wenn sowohl Mykorrhizamycel als auch Vesikel deutlich erkannt wurden. Arbuskel waren nicht in jedem Fall vorhanden. Die Liste ergänzt die Liste von Weissenhorn und Feldmann (1999, in diesem Band).
|
Abelmoschus esculentus Acacia senegal Amaranthus caudatus Anacardium occidentale Annona cherimola Anthemis tinctoria Arachis hypogaea Artocarpu heterophyllus Astragalus gummifer Aucuba japonica Bactris gasipaes Bertholletia excelsa Bixa orellana Cajanus cajan Canavalia ensiformis Cannabis sativa Carica papaya Carya illinoinensis Cicer arietinum Cichorium intybus Cinnamomum
verum Citrullus
lanatus Cocos nucifera Cola verticillata Coriandrum sativum Cuminum cyminum Curcuma longa Cydonia oblonga Cynara cardunculus Cynara scolymus Cyphomandra betacea Cyrtostachus lacka Dioscorea bulbifera Diospyros ebenum Diospyros kaki Dolichos lablab Echinocloa frumentacea Elaeis guieensis Elettaria cardamomum Eleusine coracan Enterolobium cyclocarpum Eugenia caryophyllata Eupatorium
rebaudianum Glycine max Hevea brasiliensis Hippophae rhamnoides Ipomoea
batatas Lens culinaris Leucaena leucocephala Licania rigida Luffa aegyptiaca Magnifera indica Malpighia glabra Manihot esculenta Metroxylon sagu Nephelium lappaceum Ochroma pyramidale Orbignya speciosa Paullinia cupana Pennisetum americanum Persea americana Phaseolus lunatus Pistacia vera Pisum sativum Psidium guajava Saccharum officinarum Schefflera arboricola Schizolobium amazonicum Scorzonera hispanica Sechium edule Sesamum indicum Sorghum
bicolor Sorghum
saccharatum Stephanotis
floribunda Sterculia
urens Swietenia
macrophylla Tamarindus
indica Tectona
grandis Terminalia
superba Theobroma
cacao Theobroma grandiflorum Vigna angularis Vigna mungo Vigna radiata Vigna
sinensis Vigna
unguiculata |
den Baumschulen, geringere Verluste trotz Krankheitserregern, potentielle
Einsparungen bei Pflanzenschutzmitteln, Verringerung der Auspflanzverluste nach
Ausbringung der Pflanzen ins Feld und schließlich besseres Wachstum im Feld mit
der Folge früherer Propfung mit Pathogen-resistenten Baumkronen können
ökonomisch bilanziert werden gegen einen geringen Mehraufwand an Handarbeit zum
Zeitpunkt der Inokulation (Mischung des Substrates mit dem AMP-Inokulum) und
die Kosten des Inokulums. Diese Kosten lagen inklusive der notwendigen
Investitionen (Pflanzkästen) bei maximal 10% der Kosten für erfolgreich okulierte
Jungpflanzen. Allein die Reduzierung der Auspflanzverluste machten deshalb in
diesen Versuchen den Mykorrhizaeinsatz bereits rentabel. Unsere Ergebnisse
führten dazu, daß die Berücksichtigung der Mykorrhiza im Integrierten
Pflanzenschutz des Kautschukbaumes proklamiert und umgesetzt wird (Lieberei et al., 1989; Feldmann et al., 1989; Feldmann et al, 1995).
Die Ergebnisse, die hier am Kautschukbaum dargestellt wurden, können mit
großer Sicherheit auf zahlreiche andere mehrjährige und einjährige Nutzpflanzen
der Tropen und Subtropen übertragen werden (Sieverding,
1991; Feldmann et al., 1995b;
Tab. 3). Die regelmäßige und breite Einführung der Mykorrhizatechnologie in die
Praxis der Pflanzenbauproduktion Brasiliens und von Sri Lanka allgemein
scheitert bislang vor allem an der Bereitstellung von adäquatem AMP-Inokulum
und dessen Verbreitung. Für die Bereitstellung von AMP-Inokulum muß das im Land
befindliche Know-how zentriert und in einer Anfangsphase nicht-kommerziell
organisiert werden - ähnlich wie es in der Anfangsphase der Einführung der
Mykorrhizatechnologie auch in europäischen Ländern über die Universitäten
gewährleistet war. Eine Verteilung des AMP-Inokulums und die Schulungen, damit
in geeigneter Weise umzugehen, müssen zweifellos von landwirtschaftlichen
Beratungsstellen geleistet werden.
Nicht organisatorisch, sondern biologisch begründete Schwierigkeiten bei
der Bereitstellung von Inokulum liegen in den Auswahlverfahren für die AMP für
bestimmte Kulturen. Wie oben dargestellt sind spezifische Wechselwirkungen
zwischen Pilzlinie und Pflanzensorte ein Haupthindernis für die nutzbringende
Umsetzung eines geplanten Einsatzes. Auch für die Überwindung dieser
Schwierigkeiten ist es in beiden Ländern unumgänglich, die genannte
Institutionalisierung der Inokulumproduktion und die enge Zusammenarbeit
zwischen diesen Einrichtungen und den genannten Beratungsstellen in den
Mittelpunkt der Bemühungen zu stellen. Auf diese Weise könnte das aufgezeigte
Nutzungspotential der natürlichen Symbiosen insbesondere von den
Bevölkerungsteilen ausgeschöpft werden, die sich teuren Mineraldünger nicht
leisten können. Orientiert man die Ergebnisse zur mykorrhizabedingten Zunahme
des Phosphor-Entzuges (Tab. 1) in den vorliegenden stark P-fixierenden Böden an
den Analysen von Pereira et al. (1988),
so kann eine vergleichbare Steigerung des P-Gehaltes in nicht-mykorrhizierten
Kautschukbäumen nur durch eine Erhöhung der P-Düngung von 150kg/ha auf
1200kg/ha erreicht werden. Eine Vernachlässigung der Mykorrhizatechnologie
hieße hier, sich gegen den wirtschaftlichen Anbau von Hevea zu entscheiden.
Der experimentelle Einsatz von arbuskulären Mykorrhizapilzen in der
Pflanzenproduktion des Kautschukbaumes erwies sich unter Praxisbedingungen als
ökonomisch sehr sinnvolle Pflanzenbaumaßnahme. Besseres Wachstum der
Jungpflanzen und damit kürzere Standzeiten in den Baumschulen, geringere
Pflanzenverluste in Gegenwart von Krankheitserregern, potentielle Einsparungen
bei Pflanzenschutzmitteln, Verringerung der Auspflanzverluste nach Ausbringung
der Pflanzen ins Feld und schließlich besseres Wachstum im Feld mit der Folge
früherer Propfung mit Pathogen-resistenten Baumkronen können ökonomisch
bilanziert werden gegen einen geringen Mehraufwand an Handarbeit zum Zeitpunkt
der Inokulation (Mischung des Substrates mit dem AMP-Inokulum) und die Kosten
des Inokulums. Diese Kosten lagen inklusive der notwendigen Investitionen
(Pflanzkästen) bei maximal 10% der Kosten für erfolgreich gepropfte
Jungpflanzen. Allein die Reduzierung der Auspflanzverluste machten deshalb in
diesen Versuchen den Mykorrhizaeinsatz bereits rentabel.
Teile der vorgestellten Arbeiten wurden finanziert aus Mitteln des deutschen BMBF+T über das Programm SHIFT, aus Mitteln des Institutes für Angewandte Botanik, Universität Hamburg, des CPAA/EMBRAPA, Manaus, Brasilien, des RRISL, Agalawatta, Sri Lanka, über Stipendien des Landes Niedersachsens, des DAAD und der brasilianischen CAPES.
Feldmann, F., 1991: Die Mykorrhiza des Kautschukbaumes
Hevea spec. Müell. Arg.: Vorkommen am Naturstandort, Auswirkung auf das
Resistenzverhalten, Nutzung im Plantagenbau. Dissertation, TU Braunschweig,
1991
Feldmann, F., Idczak, E. , 1994: Inoculum production of VA-mycorrhizal fungi. In: Norris, J.R.; Read, D.J., Varma, A.K. (eds.): Techniques for mycorrhizal research, Academic Press, San Diego, 799-817
Feldmann, F., Idczak, E., Martins, G., Nunes, J., Gasparotto, L., Preisinger, H., Moraes, V.H.F., Lieberei, R., 1995a: Recultivation of degraded, fallow lying areas in central Amazonia with equilibrated polycultures: Response of useful plants to inoculation with VA-mycorrhizal fungi. Angewandte Botanik, 69, 111-118
Feldmann, F., Idczak, E., Nunes, C.D.M., 1995b: A importancia dos fungos micorrizicos no manejo de sistemas agricolas na Amazonia. In: Kanashiro,M, Parotta, J.A. (eds): Manejo e reabilitacao de areas degradadas e florestas secundarias na Amazonia; USDA, Puerto Rico, USA, S. 57-65
Feldmann, F., Junqueira, N.T.V., Lieberei R., 1989: Utilization of vesicular-arbuscular mycorrhiza as a factor of integrated plant protection. Agriculture, Ecosystems and Environment, 29, 131-135
Feldmann, F., Lieberei, R., 1994: Vesicular-arbuscular mycorrhiza in rubber tree monocultures. Mycorrhiza News (New Delhi) 5 (4), 1-6
Janos, D.P., 1987: VA mycorrhizas in humid tropical ecosystems. In: Safir, G.R. (ed.): Ecophysiology of VA Mycorrhizal Plants. CRC Press, Boca Raton, Fl, USA, 107-134
Jayaratne,
A.H.R., 1982: Endomycorrhizas of rubber growing soils of Sri Lanka. J. Rubb.
Res. Inst. Sri Lanka 60, 47-57
Junqueira, N.T.V.;
Chaves, G.M.; Zambolim, L.; Alfenas, A.C., Gasparotto, L., 1988: Reacao de clones de Seringueira a
varios isolados de Microcyclus ulei. Pesq. Agropec. Bras. 23(8), 877-893
Lieberei, R., Junqueira, N.T.V., Feldmann, F., 1989: Integrated disease control in rubber plantations in South America. Proceedings of the '89 Integrated Pest Management, February 8-15, 1989, Bad Dürkheim, FRG, 445-456
Liyanage, A. de S., Dantanarayana, D.M., 1983: Association of Fusarium solani with root lesions of rubber showing leaf wilt in Sri Lanka.Trans. Br. Mycol. Soc. 80(3), 565-567
Pereira, E.B.C.; Pereira, A.V.; da Silva, S.E. L., 1988: Niveis de N, P, K, e Mg para viveiro de Seringueira em latossolo amarelo de textura muito argilosa. R. Bras. Ci. Solo 12, 143-146
Sieverding, E., 1991: Vesicular-arbuscular mycorrhiza management in tropical agrosystems. GTZ, Eschborn, Deutschland